Strategies to use on your farm: a breeder’s perspective
By Brett Grohsgal
Global climate change is real. The few citizens and elected officials who deny the heavy weight of scientific evidence may temporarily slow some societies from taking a better path, but meanwhile all agriculture must adapt very fast.
I write this article to empower farmers and crop breeders: we can and must build a better line-up of crops that will feed humans in the more difficult future.
The new normal of global climate change causes:
Less overall predictability in big weather patterns, eg., hurricanes, anomalous jet stream behavior between the US and Europe, etc. Records are now routinely broken –of typhoon wind strength, of open shipping lanes through Arctic ice, of tornados in non-tornado regions, and on and on.
Higher variability in precipitation. Our farm’s examples: we began this business in 1996. Since then, we have had the first, second, fifth, seventh and tenth worst droughts in our region since recording weather data began in 1840. We also had the wettest year on record: 2018, with 81 inches. Some Shenandoah growers got 101 inches in 2018, none of that coming from hurricanes.
Higher overall temperatures, but extremely variable. Summers are increasingly hot or bizarrely cool. Too-warm winters now trigger premature bud break in fruit trees, followed very often by one last killing frost. Radical temperature fluctuations have slowly become what we all expect.
Rising sea levels. Lucky indeed are coastal denizens who live on higher ground. Some islands are disappearing already. Salinization of low-lying coastal areas is spreading, making swathes of previously productive cropland too salty for crops.
Technology offers us an apparent alternative to weather-induced production uncertainty: greenhouses, on a massive scale. Across fruit, vegetable, and medicinal crops, and in so many nations, greenhouses and their ilk play an increasing role. The high yields of greenhouse production are a joy.
But we need to be honest with ourselves about the very heavy carbon footprints of most greenhouse production. Manufacturing their metal and plastics, cooling them in warm weather and heating in winter, pumping water instead of letting rains irrigate for free, frequent spraying to protect those valuable crops, fertilizing with exogenous nitrogen compounds instead of with green manures-all these and more shout the high carbon footprint of such protection. I argue all this as a production farmer with three simple greenhouses that hugely augment our open-field production.
As important in this era of bumpy weather, nearly all greenhouse crops depend on the reliable and uninterrupted supply of energy. When the grid goes down or the water stops running, even briefly, there is real trouble and potential widespread mortality of the foods inside.
What does this have to do with crop genetics? The answer: don’t think we can solve this by putting physical barriers between global climate change and our crops. We must not delude ourselves into believing that a techno fix like greenhouses precludes the need for crop genetic adaptation. Open-field agriculture will remain the biggest source of human foods.
These are scary truths. But witness our strengths. Anyone doing big-population breeding for at least three years (or even simple decently-executed seed saving) has already been selecting for at least some crop tolerance to climate change. Indeed, humans have been successfully breeding crops for at least 10,000 years, and this is hardly the first time that pathogens, insect pests, or climate have mandated change.
Most critical for me, the genetic libraries of many plants are amazing, diverse, and adaptive. Finally, taxpayers’ dollars have thankfully supported cold-storage seed banks in many countries. The older and more durable genelines (a term similar to variety) the seed banks steward are extensive informational reservoirs of potential resistances to the ravages of climate change.
The rest of this article covers the specific stressors that our crops will increasingly face; our genetic goals are tolerance for or resistance to these. I sprinkle illustrative breeding examples from our farm throughout the article to show both breeding successes and failures. Last, I will address the pluses and minuses of three different breeding approaches that we can use to try to cope with global climate change.

Above is one of the author’s brassica fields in flower for Recurrent Mass Selection (RMS).
The stressors
From the breeder perspective, climate change has two main paths. First, gradual upward or downward shifts in temperature or rainfall. This likely occurred with the Ice Ages, and likely is what we’re seeing with Alaska’s permafrost and glacial melting. Gradual changes are easier to breed for.
Contrast these with cataclysmic impacts of climate change, in which really ugly happens really fast. Examples include rashes of super-strong typhoons or hurricanes, or frequent summer temperature spikes to 105° F in unprepared regions. Cataclysmic is much harder to breed for. A close-to-home example for our farm were the storms Irene and Lee, only five days apart, that in total gave us 22 inches of rain in late August, our peak summer food-crop and seed-crop production period.
Higher but variable temperatures
The unpredictability of temperatures will really test many crops. Germination in abnormally cold or hot soil is challenged; dying seeds don’t make good or photo- or fertilizer-efficient stands. Perhaps we will need to accept heterogeneous germination, as we see in vetch, in which many seeds germinate quickly, but a few delay and only later germ (the so-called hard-seed phenomenon). Eventually the field gets plant coverage.
But this would be problematic for grain farmers, the vast majority of whom mechanically harvest. Vegetable growers may be able to cope with this better; starting transplants under controlled conditions is already something our sector knows well. Priming seed has also helped our operation. But until we source older genelines that are less picky with their preferred germination temperature, I doubt crop breeding will be of much help.
Flowering and successful pollination are often very dependent on both photoperiod and temperature, and on the interaction between the two. Our farm’s genetic goals for our winter greens (detailed in GFM articles in Aug.-Oct. 2002; Aug.-Sept. 2004; Feb. 2017 and more) include delayed flowering until pollinators emerge from dormancy. We want to harvest leaves, for sale, from late October until April or May, and only then see the plants flower and make seed to sow in autumn.
Decades of breeding, coupled with timing the sowing of these crops, makes this work. But one Even’ Star geneline that is vital to our production just won’t comply: our Ice-Bred Arugula. This coyote of the greens world now has not only zero mortality all winter, but it can flower during a warm spell and survive subsequent deep freezes (premature flowering followed by deep freezes causes most annual plant species to die; once they flower they are metabolically vulnerable).
I first crossed two open-pollinated arugula genelines in 1992 to wipe out possible inbreeding depression. We then used our preferred breeding approach (see “recurrent mass selection” later) every single year to breed for winter hardiness. After these 28 years of breeding, even the individual flowers don’t die unless temps get below about 18° F. That is genetic success.
Contrast this with crops much harder to breed for climatic uncertainty: apples, plums, and peaches. When these trees flower before they should because of warm winters, and subsequent mild freezes or even just frosts occur, the flowers die and are aborted. These years, too often an entire region’s tree fruit crop fails because of this. I am often thankful that arugula, not a tree fruit, is among the crops I breed.
Another crop with temperature problems are Capsicum annuum, sweet peppers of all types. In most varieties, the pollen tubes will abort if they experience temps above 92-94°F. Hence in hot summers we see low fruit set in August, and better production once cooler September weather comes. Regions further south will likely have more difficulty in raising sweet peppers as average summer temps climb. Appropriate breeding approaches to combat this would use sweet pepper genelines whose pollen tubes continue to grow at temperatures above 94° F, and cross these with peppers with market-desired traits.”
Insects
Most insects and other arthropod pests are advantaged when temperatures rise. They reproduce and do many other things faster with more heat. Higher average global temps in all seasons leads to more generations of many insect pest species per year. Higher insect metabolic rates and activities- feeding, molting, mating- must also be expected.
Shortened generation times over longer warm periods = more generations = greater insect pressure. More generations/year = more insect sex (remixing of genes), and faster adaptation to our sprays. All warming regions will experience less winter-kill of dormant arthropod pests.
Finally, the northern hemisphere will see more invasions of southern pests; flip that for southern hemisphere farmers. Most illustrative on our farm: nearly all winters we now routinely find cucumber beetles, cabbage looper larva, and even harlequin bugs- actively feeding- in our open-field Brassicaceae crops. We have not seen 6° F, a lovely temp to kill many insects, since early 2018. Right now I would be happy to get even a few nights at 15° F. I yearn for pest mortality.
Breeding for crop tolerance to increased insect pressure is difficult. Among the excellent intentional successes are the non-GMO potato varieties Prince Hairy and King Harry, whose pubescent (hairy) leaves repel Colorado potato beetles, potato leaf hoppers, etc. Incidental breeding successes include the waxier-leafed and blue-green kales, collards, broccoli, and their kin, which at least on our farm show less damage from flea beetles than do their relatives like arugula, turnips, mizuna, choi, etc.
Breeding for physical barriers to arthropod pests (like trichomes, hairs, or sclereids, the stone cells found in the fruit of pears) may be more durable than chemical means of control. Witness the myriad co-evolved insect pests of the Solanaceae (tobacco, tomato, etc.), a family that has spent eons adding nasty compounds to leaves and even flowers.

One of the author's varieties is Ice-Bred arugula, seen here in flower.
Microbial pathogens
Our operation has had greater success in breeding crop tolerances to microbial pathogens. First, the bad news about this last stressor: everything noted above about insects and rising temps applies to bacterial, fungal, and viral pests. More generations per unit time is the critical equation.
More bad news: bacterial plant pathogens can likely transfer their resistances (to plant defenses and/or to our chemical controls) through phage- or plasmid-mediated conjugation, as hospitals have seen in MRSA. Eons before human GM breeders bridged the species barriers by inserting DNA, viruses and bacteria did . . . how wonderful. Final bad news: the faster reproductive rates of most microbial pathogens gives them the capacity to co-evolve with the defenses humans generate, and often to win the co-evolution race.
We really need some good news right about now. Here goes: the genetic libraries of all of Earth’s plants and animals already contain information and directions –i.e., DNA—that can and do help these organisms fight fungi, bacteria, and viruses. All humans would soon die upon emerging from the womb if we had no immune systems.
Plants are the same. This war has been going on for billions of years, and thankfully ancient and modern breeders have had much opportunity to tap into the genetic potentials of our crops. The most effective breeding approaches appear to be targeted hybridization and/or recurrent mass selection; I know of no GMO crop varieties that as yet have successfully and durably defended against co-evolving microbes. Tolerance or resistance to pathogens will undoubtedly require multiple genes and likely multiple chromosomes and metabolic pathways (more on this later).
More good news, and supporting evidence for the above positives: our farm’s breeding work has successfully bred tolerances of or resistances to certain microbial pathogens into some of our most financially critical crops. Easiest was tobacco mosaic virus; that resistance was attained by 2002 in our best tomato genelines. Second was mosaic virus of the Brassicaceae: in our bred varieties, of the hundreds of thousands of individuals we grow every winter, only about 3 get infected/100,000, compared to about 50 per 100,000 plants in purchased brassica varieties.
Resistances to Fusarium and Verticillium wilts, both fungal, were easy in our core tomato genelines. Genetic work in progress includes our fight against Septoria, with decent to good results. Still winning against us mere humans is Xanthomonas. Although two of our genelines show a little promise, Xantho humbles me. But then I am too enslaved to production farming, unfortunately too busy, to allocate even 6 hours weekly to crop breeding. Imagine what 5 good labs and their crews could accomplish. Success correlates with directed efforts and time spent.
Breeding to manage risk: yield timing
The above outlines some of the stressors, the things we need to select for or against in future crops. But thinking more like a production farmer than like a breeder also needs to occur. One way to categorize food crops in the face of global climate change is traditional: the determinate vs. the indeterminate varieties. How acute is a crop’s harvesting bell curve? What will our yield be per unit time, and when will harvests end and the crop just die? A good way to assess determinate vs. indeterminate genelines are residence times in the field and the interaction of those times with probabilities for bad weather or increased pest pressures.
The ideal determinate crops reliably produce a lot in a short period, and then die off as predicted. Their critical period of vulnerability is short, and yields will very likely be high. Only really bad weather in that brief vulnerable period usually causes failure, and it will likely be total.
Roma tomatoes and hybrid squash in summer, and Shanghai pac choi in winter, are these for our farm. Once they are start their intense yield periods, the determinate crops cause me less worry in the middle of the night. Breeding for these “live fast, pump out lots of food fast, then die” crops is a worthwhile goal, although not a path our farm has taken: we buy those seeds.
Indeterminate crops are very different. Here the plants must live longer, but they can survive periods of bad weather and then, eventually, reliably produce a decent (or better) crop. Their high residence time (= extended period of vulnerability) is offset by their resilience to much of the nastiness that our changing climate can throw at them.
Ideally the breeder also selects for early, middle, and late production in the indeterminate crops. Certainly, our farm has found it very simple to select for both early and extended yields in our tomato genelines and some peppers. Less simple was selecting for tolerance to microbial pests. But this crop reliability was purchased with decades of our farm’s genetic labor.
Worst case crops in terms of risk management include modern corn and modern strawberries. Modern corn, most genelines of which are hybrid and/or GMO, has both a lengthy residence time and at least 3 periods of high vulnerability: planting window with soil temperatures; at silking; and close to harvest, when (as in nearly all crops) excess rain or hurricanes can doom yields.
Some older corn varieties staggered their flowering, with 3-plus potential ears. Our farm no longer grows corn. But we do unwisely grow commercial strawberries, a super-popular crop with profound vulnerability to excess rain for the five weeks of fruit ripening.

Another one of the varieties the author bred, Chinese Thick-Stem mustard. It and Ice-Bred arugula are available from Fedco Seeds.
Three breeding approaches
This is a simplified discussion in the interests of your time. For more depth, please refer to my 2002 articles, Seed saving part one, two and three, in the Aug-Oct Growing for Market magazines. Here are three basic ways to breed crops.
Hybridization/pedigree line programs: these traditionally start with 2 inbred lines and small, relatively unrelated parent populations. The F1 offspring are remarkably uniform. An easy example is crossing two Cucurbita pepo squash open-pollinated cultivars. But perhaps the most promising hybrid programs do not start with inbred lines, but instead with more diverse open-pollinated genelines, as did (separately) our Ice-Bred Arugula and our New Star Mustard. We attained strong hybrid vigor (=heterosis) in both new landraces, in both adapted genelines. Heterosis does not appear to be strictly owned by inbred isolated genelines, nor should skilled breeders limit their tools to two inbred homozygous lines.
GMO approaches: typically depend on a mutation of a single gene, or very few linked genes, or insertion of valued foreign DNA into the crop’s genetic code. Recently some GM breeders have attained stacked tolerances, almost as good as what RMS (see below) routinely accomplishes. But inheritance of complex and multi-chromosomal metabolic pathways remains mostly a dream.
Recurrent mass selection (RMS): depends on large populations of potential breeders (mass), many or all of which should be genetically diverse. Harsh screens are imposed before pollination and sex begin (selection). Death is good. Survivors are permitted to make seed. The fruit or mother plants are GROUP or INDIVIDUALLY selected to make the next generation. Ideally the crop is not babied; ideally RMS happens every year (recurrent); ideally first-year death rates are high but the breeder still has 50-2000 survivors for the screened next population (again, mass).
Now let’s assess the pluses and minuses of the 3 breeding approaches for their abilities to make crops resilient to climate change.
Hybrid breeding: Since traditional hybridization starts with small populations of two inbred lines for the original cross, this is a built-in minus, due to low genetic diversity within each inbred line. We see this in our fields: many hybrids are prima donnas; field conditions, fertility, the weather—all must be perfect for outstanding growth and yields.
A big plus to this breeding approach is hybrid vigor. Heterosis is real and powerful and very often profitable, but perhaps oversold by seed companies who want you to buy. See discussion in the must-read First the Seed, by J.R. Kloppenburg.
Another plus of many hybrids is early yield, which decreases the residence time (and hence vulnerability) of crops in the field. Another counterargument in favor of targeted hybridization: about half of the super-adapted, durable landraces our farm has bred have started by creating hybrid(s). Only thereafter did we then develop the superior landrace from that F1 generation.
The minuses of GM approaches in the face of global climate change revolve around the difficulties in adding whole new metabolic pathways, and the many genes usually involved, into a crop. GM is good at inserting just a few genes, or inducing mutation in one gene, but drought tolerance or salt tolerance or the abilities to co-evolve with rapidly-reproducing pathogens are just three cases in which many plant genes must be expressed properly and in coordination.
Recurrent mass selection mimics and even accelerates the billions of years of evolution that life on Earth has endured. Its minuses are that large initial populations are needed. That requires more seed, space, and likely money for the grow-outs. A second big minus is that for RMS to work right, the breeder (or farmer) must see some individual plants fail to reproduce, or better yet die. Finally, flowering (genetic exchange) must happen at about the same time within the whole surviving population. But now the pluses:
RMS shares with evolution the reliance on a genetically diverse population. That informational diversity within large and heterogeneous populations is hugely useful when a breeder, like evolution, is trying to make a species survive different stressors that all attack at the same time. At the chromosomal and loci level, RMS is by far the most multi-variate of the breeding approaches.
Once you are a few generations into the land race, the acceptable and indeed desired mortality of certain percentages of the total breeding population need only be about 20-30% for RMS to continue to improve the landrace. Farmers can still break even or make some money if losing 30% of a stand, especially if the stronger surviving plants take advantage of the newly-opened space and fertilizer. That mortality likely decreases every year.
If each year’s crop has progressively lower mortality, the future is more promising than when 100% of some prima donna crop dies. An example is our Chinese Thick-Stem Mustard. Two provenances were pathetic the first 2 years of attempted breeding: those two lots died at normal temperatures below 22° F. Only the pre-deep-freeze harvests generated income.
In year #3, I tried a third provenance that had 70% mortality over an entire winter. The 30% that survived got to have sex, once spring arrived, and those seed were then planted the next year. Every year the previous winter’s survivors get to breed. By now, 19 years later, mortality has been zero even in that rare winter that saw our temperature briefly dip to -1° F. A green that is definitely delicious, definitely profitable.
Recurrent mass selection does not require a lab, nor a team of pollinating (or de-tasseling) humans. Oversimplifying, maybe: let the bad happen, and as long as the starting population is big enough, the breeder/farmer has good chances of success.
By not fearing environmental stressors but instead embracing them, RMS is the most nimble of the breeding approaches. Yes, it is the harshest of the breeding approaches, the most Spartan. But then so is climatic uncertainty, our new normal.
Earlier I noted the 22 inches of rain our farm got over only five days from Hurricane Irene and Tropical Storm Lee . . . a horrible August indeed. But the five Even’ Star proprietary cherry tomato genelines and the bigger heirloom Cherokee Purple that survived and then ripened fruit were gold. Not because of easy sales, although rarely has the mid-Atlantic seen tomato prices so high.
No, it was the seed within the survivors that made them so valuable: those plants withstood 22 inches of mostly salty rain (we are only 1500’ from the Chesapeake, so storms like those produce saltier downpours than gentler events). Those plants also withstood the myriad fungal and bacterial pathogens that became epidemic that particular September. Perhaps most important, that cataclysm was only one in a long series of stressors that had tested and continue to test the genelines.
Ideally for me, we humans would be honest with ourselves. We would admit that we are in the midst of global climate change. Only then can we take wise steps to survive. Breeding crops- the plants that will feed us tomorrow, in ten years, in fifty years- is the part of adapting to this new normal that I want to take on. May you, too, join me in breeding better crops for our future.
This article is dedicated to Cooperative Extension agents and to the ag departments of our universities. They need our support and will pay back our investments many times over. As well, I deeply thank the countless forgotten farmers and breeders who, over the millennia, developed nearly all the crops that feed us now. We will survive. And we will continue those farmers’ work.
Brett Grohsgal crossed his first crops in 1992, has commercially farmed since 1996, and has periodically written for GFM on crop genetics, winter cropping, farm risk management, buying farmland, etc. He and his wife Christine Bergmark own and manage the 104 acres of Even’ Star Organic Farm and market their crops through summer and winter CSAs, farmers markets, and wholesale.


 In the April 2020 issue, Brett Grohsgal’s article “
In the April 2020 issue, Brett Grohsgal’s article “
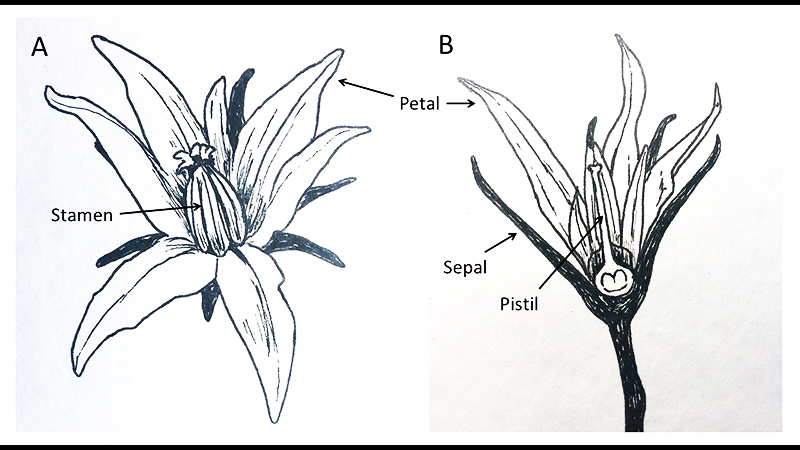 In the April 2020 issue of Growing For Market, I focused on the selection side of tomato breeding. In this follow-up article, I will discuss how to make crosses and manage breeding projects.
In the April 2020 issue of Growing For Market, I focused on the selection side of tomato breeding. In this follow-up article, I will discuss how to make crosses and manage breeding projects.
 Farmer to Farmer Profile
Farmer to Farmer Profile
 I met up with Fred Hempel, the Artisan Seeds breeder, at his farm in Sunol, California on Sunday, October 8, 2017. It was a bright, dry 77-degree day, typical for east of the San Francisco Bay Area in the fall. It would have been even brighter if it weren’t for the thin layer of dust covering everything, dimming the reflection from the sun. In testament to how dry it was, historically bad wildfires would break out later that evening farther north in Sonoma County and burn many acres of farmland, vineyards and homes over the following days.
I met up with Fred Hempel, the Artisan Seeds breeder, at his farm in Sunol, California on Sunday, October 8, 2017. It was a bright, dry 77-degree day, typical for east of the San Francisco Bay Area in the fall. It would have been even brighter if it weren’t for the thin layer of dust covering everything, dimming the reflection from the sun. In testament to how dry it was, historically bad wildfires would break out later that evening farther north in Sonoma County and burn many acres of farmland, vineyards and homes over the following days.



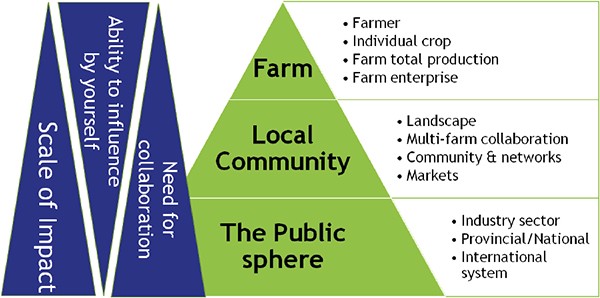 While lobbyists and some politicians have managed to paint the climate change debate as a partisan issue, farmers are increasingly feeling the impacts on their production environment and asking themselves: How can I re-think my operation in order to continue farming with increasingly unpredictable weather conditions? In this article, I am summarizing my lifelong quest for understanding climate resilience and offer a self-assessment template to help you plan for more resilience.
While lobbyists and some politicians have managed to paint the climate change debate as a partisan issue, farmers are increasingly feeling the impacts on their production environment and asking themselves: How can I re-think my operation in order to continue farming with increasingly unpredictable weather conditions? In this article, I am summarizing my lifelong quest for understanding climate resilience and offer a self-assessment template to help you plan for more resilience. 










