By Fred Hempel
In the April 2020 issue of Growing For Market, I focused on the selection side of tomato breeding. In this follow-up article, I will discuss how to make crosses and manage breeding projects.
The nuts and bolts of crossing tomatoes
Crossing two different tomato varieties can be done by physically moving pollen from the flower of one variety to the flower of another. This is accomplished in a series of steps. First, one variety is chosen to be the mother and another the father. I like to use smaller-fruited varieties as mothers. This seems counterintuitive, but from experience I have found that large beefsteaks produce fewer seeds per fruit and take longer to ripen.
Since tomatoes have male and female reproductive structures on the same plant, any tomato plant could be a mother and/or a father. In fact, tomatoes have perfect flowers (i.e. male and female parts in the same flower), and typically function as inbreeders (self-crossers). If left to their own devices, tomatoes will self-pollinate. Which is why it is most likely if you save the seeds from on open-pollinated tomato, that it will look just like the parent plant because it self-pollinated.
The likelihood of self-pollination is why the next step in the process is necessary if you want to ensure a cross with a different variety. Since each flower has male and female reproductive parts, to prevent self-pollination you must remove the male parts from the flower intended to be the mother, a process known as emasculation.

Figure 1. A complex inflorescence of tomato flowers. The opened flower with the yellow anther cone and yellow petals is at the pollen producing stage. Pollen is being shed inside the cone. The flowers labeled “1” and “2” are at the right stage for emasculation and cross-pollination. Although pollen is not shedding in these flowers, the stigmatic surface (not visible) is receptive to pollen. After emasculation (by removal of sepals, petals and the anther cone) the pollen can be brushed across the receptive stigma. Once pollination has been completed, a small pair of scissors should be used to remove all other flowers and buds in the photo. Image courtesy of Dan Waber.
Once a mother plant is chosen, I use jeweler’s tweezers to remove the floral structures from a few of the flowers on the mother plant. The flowers to be emasculated are the ones with light yellow petals that have not yet unfolded. In other words, they are still tightly appressed to the anthers in the flower bud (see figure 1 above). The oldest unopened flowers are appropriate for emasculation because they have not yet begun to shed pollen and still have a stigma that can receive pollen (see drawings below).
Working from the outside inward, I remove the sepals, the petals, and finally, the cone of anthers (pollen-bearing structures) that surround the ovary/style/stigma (female structures). I have found that removing the sepals and petals at the base first makes it much more likely that the anther cone can be removed as a single unit, thus reducing potential damage to the stigma and style. The removal of the sepals is also useful because it provides an indicator for hand-pollination in case the tags fall off or are ambiguously placed.
The general process of tomato flower emasculation and pollination is demonstrated in a number of YouTube videos available online.
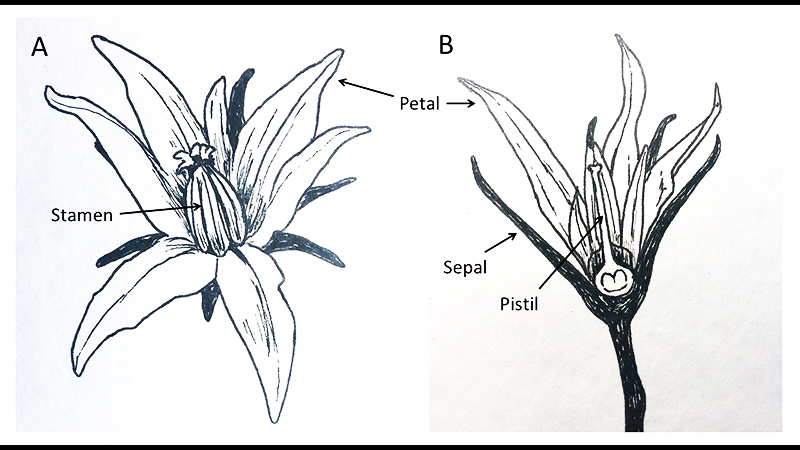
Tomato flower structures. In the complete flower (A) all structures are visible and the stamens completely enclose (and obscure) the pistil. Sepals are the outermost “whorl” of structures, and the petals, stamens and ovary/pistil are arranged from outside-to-inside the flower (B). Drawing by Nicole Hempel.
After I have emasculated a few flowers, leaving only the ovary/style/stigma intact, I remove all of the older flowers and also all of the younger flowers on the same branches as the emasculated flowers. This accomplishes two things. First, removing all nearby flowers makes it easier to tag and follow exactly which flowers have been crossed. Removing nearby flowers also reduces the competition for resources by other developing fruits in that region of the plant.
Since hand-crossing is typically less efficient than allowing the plant to self-cross, the fruits made by hand-crosses produce fewer seeds. In turn, they are less efficient at early stages of fruit ripening that signal the plant to “feed” the fruit. If other fruits are left in the region, they can suppress the growth of the fruits developing from hand-crosses.
Identifying project goals
The reason for clearly defining project goals is simple: It is better to put extra work into developing a few exceptional new varieties as opposed to creating a large number of new mediocre varieties. The challenge in breeding is not to make new varieties, it is to make great varieties. This is particularly true for tomato breeding since there are already many good varieties to choose from. In fact, I would argue the world doesn’t really need nor crave more “good” varieties.
For beginning breeders, I suggest starting the first year by making no more than three crosses. The parents of the crosses should be tomato varieties you love. If you use tomato varieties you grow every year and love to eat, there is almost no way you can produce “bad” tomatoes.
On the other hand, if you start out trying to fix flawed varieties, your project will likely include many mediocre tomatoes. Starting small also allows you to get a very clear handle on the process of selection. You will be less tempted to grow populations that are too small to allow you to see rare and exceptional trait combinations.
Starting small gives you the flexibility to work with unanticipated trait combinations. Although starting with defined goals helps narrow your focus, it is still very valuable to be able to recognize, follow and develop new projects that come out of the unexpected lines that you will see when you start with F2 and F3 populations large enough to produce as many trait combinations as possible. Your ability to follow up on unexpected, but sometimes uniquely valuable lines depends on starting with projects that are initially limited in size and not overwhelming.

This is the author’s new tomato Agi Red, which is an example of a rin/+ (heterozygous) hybrid tomato with long shelf life and excellent flavor and texture. Photo courtesy of Pascal Antigny.
Crossing to generate new inbred (OP) lines
We make crosses in order to create novel inbred lines. These lines are also called “true-breeding” or “open-pollinated” and they require the selection and saving of seed from self-pollinated individuals for seven generations. That is approximately how long it takes for the vast majority of the genes in a line to become homozygous. Sometimes it takes a little longer. After seven generations, continuously self-pollinated tomato lines usually produce offspring that are pretty much identical to the parent.
One reason it is relatively easy to breed tomatoes is that they preferentially self-pollinate because the cone of anthers that produce the pollen surrounds the female stigma where the pollen is deposited. This is less true for the first flowers to form on the plant, that are typically more open at the time of pollination. For this reason, collect seeds from fruits that form from flowers where the anthers completely enclose the female structures.
The new inbred lines created through crossing and subsequent rounds of selection can be used either as finished products (true-breeding or OP lines), or they also can be used as parents to make hybrids which are “finished” varieties. In this case, the crosses are used to produce hybrids that can be evaluated and used immediately without further selection.
If your primary goal is to make crosses and then to select them for seven generations to create true-breeding lines, there are some traits that seem to be relatively amenable to optimization in true-breeding lines. In our experience, flavors and colors are two traits that can be optimized in true breeding lines.
Unique plant habits (for example, dwarfing) are also relatively easy to optimize in projects designed to produce inbred lines. Therefore, these traits are good ones to focus on when your goal is a true-breeding/open-pollinated line.
Keeping track of breeding information
If you start out with small breeding projects, it is more likely you will develop good methods for organizing projects before you are overrun with data. I record and organize information related to my tomato breeding in two ways. I record both pedigree information along with some trait information on the #1 coin envelopes that I use to store seed (see photos). I also organize projects and crosses in Excel spreadsheets. These two general methods overlap, and I find that I am constantly adjusting the ways in which I organize breeding information.
My seed envelopes and the boxes they are stored in represent a flexible way to both store and organize breeding projects. Each seed envelope contains seeds typically collected from one plant. The first seed envelopes in a breeding project are those that contain the original hybrid seed, that is, the seed that was produced by the fruits that developed from crossed female flowers.
The envelope containing the hybrid seed is labeled “Variety A x Variety B” with the female parent always listed first. Below that I add the date the seed was collected. In this generation there are generally no notes below the date because the fruits producing the hybrid seed look like those of the female parent (See Figure 2a).

Figure 2. An example of seed packet labeling for breeding projects includes pedigree and trait information. Images courtesy of the author unless otherwise noted.
The envelope corresponding to the next generation (containing the F2 seed) is labeled “Variety A x Variety B” with a circled “X” below the original cross. This circled “X” indicates that the seed is the result of the self-pollinated fruits on a hybrid plant. I again include a date and below the date I include data on traits of interest (See Figure 2b). In the hybrid (F1) generation, all of the plants should look similar because they all received half of their genetic information from each parent.
The envelopes corresponding to the next generation and the F3 seeds typically contain more information related to the F2 plants. The pedigree at the top of the envelope now includes a number that indicates which F2 plant was harvested (Fig. 2c). In this case the “-1” indicates that the F1 generation seed came from the first (and typically only) plant harvested. The “-3” indicates that the envelope contains seed from the third plant harvested in the F2 generation. The traits listed (red cherry, long shelf life and flavor score) are typically the most important traits. Other traits will be listed in a spreadsheet corresponding to the field analysis for any given year.
The pedigree on the envelope containing F4 seed (Fig. 2d) indicates that the seed was harvested from the second plant that was grown from the F3 seed from the packet in Figure 2c. The notes on the seed packet also indicate that traits, in this case color, continue to segregate well after the F2 generation.
Because multiple plants can be selected from the F2 generation onward, projects can expand at each generation. All lines collected in the previous generation can produce multiple lines in the next generation. To keep projects under control, I try to not select more than four or five lines with every generation for each original cross. Thus, some lines become dead-ends.
One way to organize all of the envelopes/seeds from a cross is to store them in a small box. Figure 3 shows one of the small plastic boxes that I use to store seeds. In this case, the hybrid seed is in the front of the box and each successive generation is stored behind the generation preceding it.

Figure 3. An example of project storage. The first generation (F1 seed) is stored at the front of the box with each successive generation stored behind it.
Since envelopes are only large enough for a small number of notes, I use spreadsheets to supplement the data on the envelopes. Spreadsheets can be as simple or complex as is necessary, but my personal preference is to quickly and qualitatively score a number for the most important traits. I use a simple 1-5 scoring system for flavor, fruit color, plant vigor, disease resistance and fruit production. By keeping scoring simple, it increases the likelihood that I will actually look at the score and select all the important plants in my field.
One final comment on project organization: It is very easy to collect and save seeds from too many plants at the end of each growing season. Even a small number of breeding projects can quickly become overwhelming if we attempt to grow out all lines available. Thus, one of the most important decisions we have to make is when to drop or back-burner lines with limited potential. Remember each additional line grown likely reduces the opportunities to adequately evaluate and select from our best lines in each generation.
Crossing for F1 synergy
For years, we used crossing exclusively as a way to select for new true-breeding lines. However, our breeding approach has evolved over time to embrace the creation of hybrids as useful new varieties beyond the initial vehicle for producing new true-breeding varieties. During our years of growing and observing hybrid tomatoes, true-breeding tomatoes and tomatoes at every step in between, we saw that general hybrid vigor as well as hybrid trait synergy represent unique opportunities in the development of new varieties.
While we continue to release varieties that are true-breeding, we are also actively making and evaluating hybrid lines with the purpose of releasing them as distinct F1 varieties. In some cases hybrids offer unique opportunities that simply can’t be duplicated with true-breeding varieties.
For example, we are using the rin trait to extend fruit shelf life significantly, typically with little or no negative effect on texture or flavor. Rin is a naturally occurring trait in tomatoes, short for “ripening inhibitor,” that delays the softening of a tomato, though ripe flavors continue to develop normally. For more on rin, see the article “The man behind the tomatoes” from the February 2018 GFM.
However, the rin gene that confers extended shelf life is most effective when only one parent carries it. In other words, the rin heterozygote (rin/+) has the best shelf life/flavor characteristics. This is a clear case of hybrid trait synergy, as fruit qualities in rin heterozygotes are superior to either parent. Our new tomato “Agi Red” is an example of a rin/+ hybrid tomato with long shelf life and excellent flavor and texture.
Other traits we have found to be easier to optimize in hybrids (and very difficult to optimize in true-breeding lines) are disease resistance and plant architecture traits. In fact, the balance of leaf and fruit production in most of our true-breeding parent lines is skewed toward the overproduction of leaves, or toward the underproduction of leaves.
In the first case, the plant spends energy producing more leaves than needed to fill the fruits, in other words, it could have produced more fruits if it produced fewer leaves. In the second case, we also have parent lines that do not produce enough leaves to fill all of the flowers and fruits that they make.
Our new hybrid tomato Green Bee is an example where one parent produces leaves at the expense of flowers and fruits and the other parent is a short plant that produces many flowers and fruits early in development. Eventually the leaves cannot keep up with all of the flowers produced and the plant ceases production of fruits. However, in the Green Bee hybrid the balance of leaves and fruits is much more optimized. Significantly, more fruits are produced by the hybrid than by either parent.
In addition to trait synergy, we have also consistently observed the general phenomenon known as hybrid vigor in tomatoes. It is not surprising since whenever you create a hybrid, it has many more potential genes in the toolkit than either parent because each parent donates alternative gene forms for many genes.
Currently at our farm, we plant parental lines in the rows next to the rows containing the hybrids made from those same parental lines. In most cases, the plants in the hybrid rows are distinctly larger, more vigorous and produce more fruit compared with the plants in the “parental” rows next to them. These differences are qualitative and very obvious. No measurements are needed to demonstrate the differences; they are that apparent. The numbers of crates of fruits coming off the hybrid rows is almost always significantly greater.
There are exceptions. Many years the OP variety “Captain Lucky” could be mistaken for a hybrid from afar in our fields. The plants are large, productive and disease resistant. We also have a couple of cherry parents that seem to be as productive as our hybrid cherry varieties. But these exceptions only prove the rule that there seems to be a growth/vigor cost that accrues as you go through the de-hybridization process from hybrid to true-breeding.
Fred Hempel is a first generation farmer. He grows produce and breeds tomatoes on a small farm (Green Bee Farm) in Sunol, CA. Tomatoes he develops are sold via Artisan Seeds (www.growartisan.com) and Johnny’s Selected Seeds.


 In the April 2020 issue, Brett Grohsgal’s article “
In the April 2020 issue, Brett Grohsgal’s article “
 Strategies to use on your farm: a breeder’s perspective
Strategies to use on your farm: a breeder’s perspective
 Farmer to Farmer Profile
Farmer to Farmer Profile
 I met up with Fred Hempel, the Artisan Seeds breeder, at his farm in Sunol, California on Sunday, October 8, 2017. It was a bright, dry 77-degree day, typical for east of the San Francisco Bay Area in the fall. It would have been even brighter if it weren’t for the thin layer of dust covering everything, dimming the reflection from the sun. In testament to how dry it was, historically bad wildfires would break out later that evening farther north in Sonoma County and burn many acres of farmland, vineyards and homes over the following days.
I met up with Fred Hempel, the Artisan Seeds breeder, at his farm in Sunol, California on Sunday, October 8, 2017. It was a bright, dry 77-degree day, typical for east of the San Francisco Bay Area in the fall. It would have been even brighter if it weren’t for the thin layer of dust covering everything, dimming the reflection from the sun. In testament to how dry it was, historically bad wildfires would break out later that evening farther north in Sonoma County and burn many acres of farmland, vineyards and homes over the following days.
